




In
the Alber lab, we explore the innumerable ways in which microbes are
able to metabolize an amazing variety of carbon sources.
Using the well-studied anoxygenic phototrophic bacterium Rhodobacter
sphaeroides as a model organism, we work to uncover
properties of unknown enzymatic pathways for carbon assimilation.
The premise of our work hinges on the assertion that all
cells must be able to synthesize precursor metabolites (e.g.
oxaloacetate, pyruvate, alpha-ketoglutarate, acetyl-CoA, etc.) from
carbon substrates, a process known as anaplerosis. The recent
realization of the immense diversity of anaplerotic reaction sequences
drives us to pursue yet undiscovered pathways of carbon assimilation
while also seeking a better understanding of currently known pathways.

Growth on substrates that are exclusively metabolized via acetyl-CoA requires one of many specialized pathways for anaplerosis.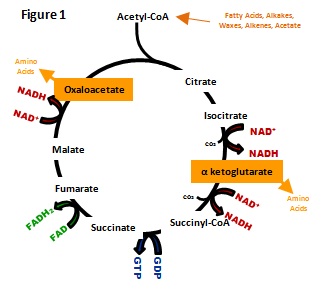 In
order to understand why anaplerotic reactions are required, one must
examine
the relationship between acetyl-CoA and the tricarboxylic acid (TCA)
cycle (Figure 1). Figure 1 illustrates that acetyl-CoA is the
sole source of carbon input for the TCA cycle while intermediates of
the cycle are necessarily removed for biosynthetic purposes.
In the absence of an ancillary pathway for anaplerosis,
acetyl-CoA, a two-carbon molecule, enters the citric acid cycle
as the sole input of carbon. The cycle outputs two carbons as CO2
while intermediates of the cycle are simultaneously withdrawn as
biosynthetic presursors. Therefore, intermediates required for
maintaining the cycle are removed from without any net carbon
input, collapsing the cycle. Ultimately, the cell needs a method
to
synthesize additional intermediates from acetyl-CoA to continue
production of the biosynthetic precursors within the cycle.
In
order to understand why anaplerotic reactions are required, one must
examine
the relationship between acetyl-CoA and the tricarboxylic acid (TCA)
cycle (Figure 1). Figure 1 illustrates that acetyl-CoA is the
sole source of carbon input for the TCA cycle while intermediates of
the cycle are necessarily removed for biosynthetic purposes.
In the absence of an ancillary pathway for anaplerosis,
acetyl-CoA, a two-carbon molecule, enters the citric acid cycle
as the sole input of carbon. The cycle outputs two carbons as CO2
while intermediates of the cycle are simultaneously withdrawn as
biosynthetic presursors. Therefore, intermediates required for
maintaining the cycle are removed from without any net carbon
input, collapsing the cycle. Ultimately, the cell needs a method
to
synthesize additional intermediates from acetyl-CoA to continue
production of the biosynthetic precursors within the cycle.
The necessity for anaplerotic reactions was immediately apparent to Kornberg and Krebbs in 1957 when th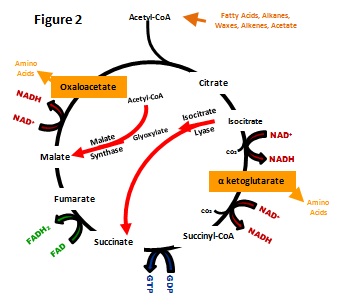 ey
identified the enzyme activities for isocitrate lyase and malate
synthase in
Pseudomonas
and Escherichia coli
(Kornberg and Krebbs, 1957). Together, these enzymes
provided a method for incorporating an additional acetyl-CoA unit while
bypassing
the CO2 releasing steps of the TCA cycle. They
had seemingly solved the question of acetyl-CoA assimilation.
With the newly
understood "glyoxylate bypass," as they termed the pathway, an organism
could assimilate carbon during growth solely on substrates that are
metabolized via acetyl-CoA. However, they shortly realized a
new complexity. Some organisms, including R. sphaeroides, do
not exhibit isocitrate lyase activity in cell extract but were fully
capable of
assimilating acetyl-CoA (Kornberg and Lacelles, 1960). For
nearly half a century, a hole loomed in the knowledge of
central carbon metabolism. No researcher was able to identify the
pathway that made it possible for these isocitrate lyase-negative
organisms to assimilate acetyl-CoA until 2007. In 2007, a
unique series of enzymatic activites was discovered in R. sphaeroides
cell
extract that
would make it possible for the organism to assimilate acetyl-CoA (Erb
et al., 2007). This series of reactions has come to be known
as the ethylmalonyl-CoA pathway (Figure 3).
ey
identified the enzyme activities for isocitrate lyase and malate
synthase in
Pseudomonas
and Escherichia coli
(Kornberg and Krebbs, 1957). Together, these enzymes
provided a method for incorporating an additional acetyl-CoA unit while
bypassing
the CO2 releasing steps of the TCA cycle. They
had seemingly solved the question of acetyl-CoA assimilation.
With the newly
understood "glyoxylate bypass," as they termed the pathway, an organism
could assimilate carbon during growth solely on substrates that are
metabolized via acetyl-CoA. However, they shortly realized a
new complexity. Some organisms, including R. sphaeroides, do
not exhibit isocitrate lyase activity in cell extract but were fully
capable of
assimilating acetyl-CoA (Kornberg and Lacelles, 1960). For
nearly half a century, a hole loomed in the knowledge of
central carbon metabolism. No researcher was able to identify the
pathway that made it possible for these isocitrate lyase-negative
organisms to assimilate acetyl-CoA until 2007. In 2007, a
unique series of enzymatic activites was discovered in R. sphaeroides
cell
extract that
would make it possible for the organism to assimilate acetyl-CoA (Erb
et al., 2007). This series of reactions has come to be known
as the ethylmalonyl-CoA pathway (Figure 3).
At its essence, the ethylmalonyl-CoA pathway incorporates three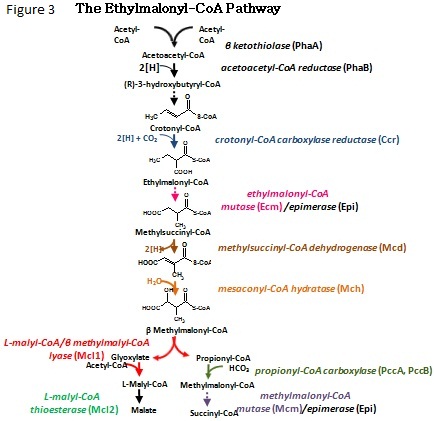 acetyl-CoA
units and two CO2
equivalents and produces
succinyl-CoA
and malate, two citric acid cycle
intermediates. The first step is the condensation of two
acetyl-CoA molecules to
form
acetoactyl-CoA. After a series of hydrations, carboxylations,
and carbon rearrangments, the pathway passes through its
namesake, ethylmalonyl-CoA, which is converted to beta-methylmalonyl-CoA.
The latter is then cleaved to form two branches in the pathway.
One branch leads
to malate by incorporation of a third acetyl-CoA unit while the other
branch proceeds through a set of reactions known from
propionate metabolism to form succinyl-CoA. All of this is
done without any loss of carbon to
carbon dioxide. In fact there are actually two carbon
dioxide units
that are co-assimilated.
acetyl-CoA
units and two CO2
equivalents and produces
succinyl-CoA
and malate, two citric acid cycle
intermediates. The first step is the condensation of two
acetyl-CoA molecules to
form
acetoactyl-CoA. After a series of hydrations, carboxylations,
and carbon rearrangments, the pathway passes through its
namesake, ethylmalonyl-CoA, which is converted to beta-methylmalonyl-CoA.
The latter is then cleaved to form two branches in the pathway.
One branch leads
to malate by incorporation of a third acetyl-CoA unit while the other
branch proceeds through a set of reactions known from
propionate metabolism to form succinyl-CoA. All of this is
done without any loss of carbon to
carbon dioxide. In fact there are actually two carbon
dioxide units
that are co-assimilated.
With the enzymatic reaction series elucidated, our next task is to study the regulation of the ethylmalonyl-CoA pathway. We are currently seeking to identify the level of regulation that is primarily used by R. sphaeroides to control the expression of this pathway and which genetic elements and gene products might be involved in regulating the pathway.

The C3 compound 3-hydroxypropionate (CH2OH-CH2-COO−) is increasingly being recognized as an important intermediate or end product of carbon metabolism in a variety of organisms. So far, there are at least five known metabolic processes involving 3-hydroxypropionate.
1. Propionyl-CoA metabolism in plants:
2. Some autotrophic CO2 fixation pathways:
3. Dimethylsulfonopropionate (DMSP) metabolism.
4. Uracil degradation:
5. Anaerobic metabolism of glycerol:
Considering its abundance, 3-hydroxypropionate is likely to play an important role in the overall carbon cycle as an end product or intermediate in the carbon metabolism of a variety of compounds. However, its use as a carbon source in bacteria is poorly understood.
Figure 4
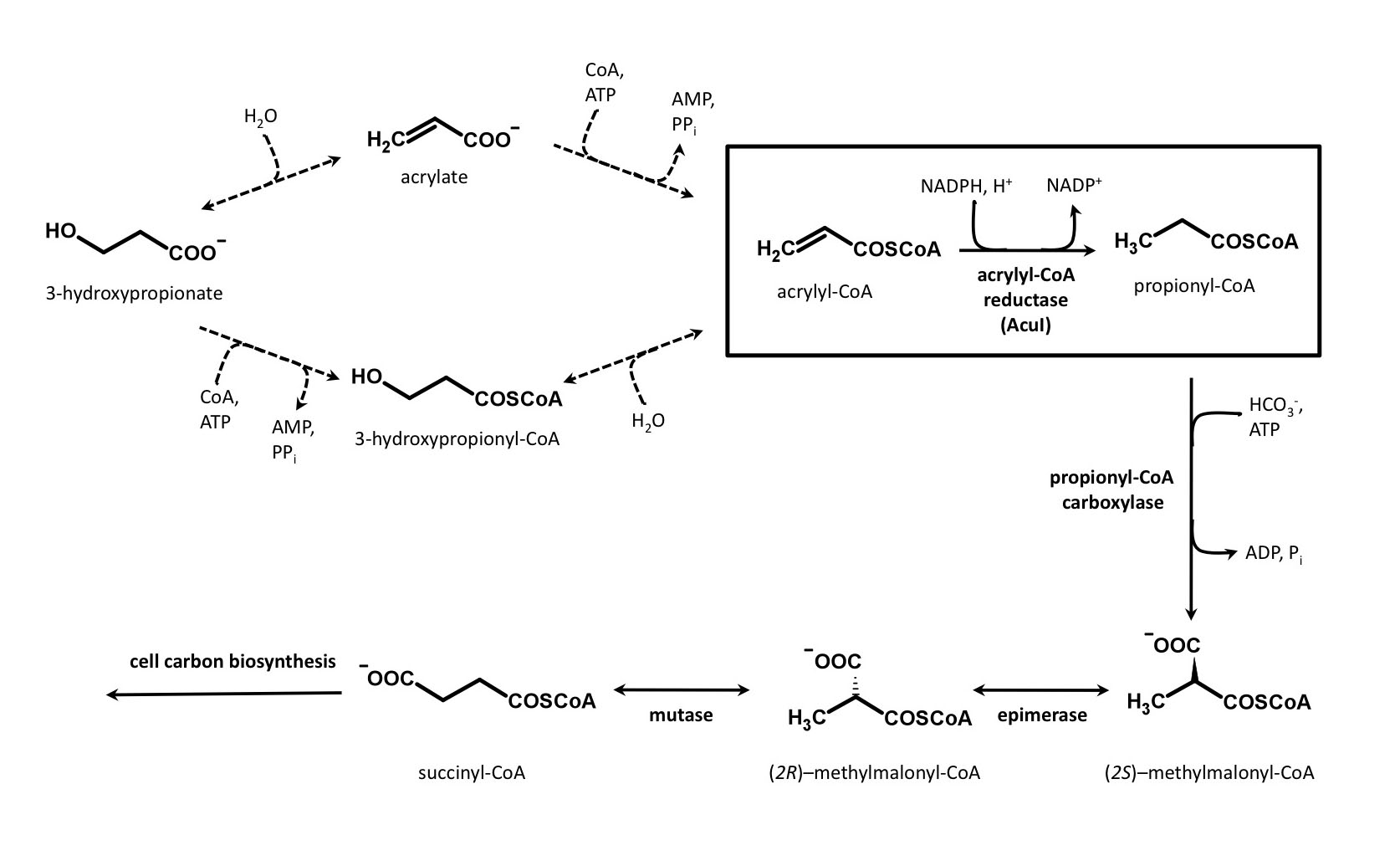 Rhodobacter sphaeroides
likely encounters 3-hydroxypropionate in its environment, as this
compound is released from other organisms. In addition, we have
demonstrated that R. sphaeroides is
able to use 3-hydroxypropionate as a sole carbon source (Schneider,
2011). If 3-hydroxypropionate were exclusively oxidized to
acetyl-CoA and CO2, a functional ethylmalonyl-CoA pathway for
acetyl-CoA assimilation would be essential to convert acetyl-CoA for anaplerosis (see above).
However, inactivation of the gene (ccr) for crotonyl-CoA carboxylase, a neccessary enzyme of the ethylmalonyl-CoA pathway, did not affect growth during 3-hydroxypropionate-dependent growth. Instead, acuI,
encoding acrylyl-CoA reductase (Asao and Alber, 2013), was
shown to be essential for growth with 3-hydroxypropionate and for the
reductive conversion of 3-hydroxypropionate to propionyl-CoA in cell
extracts. Upon production of propionyl-CoA, the cells would be
able to generate succinyl-CoA from which all of the cell's precursor
metabolites can be manufactured (See Figure 4), obviating the need for the ethylmalonyl-CoA pathway.
Rhodobacter sphaeroides
likely encounters 3-hydroxypropionate in its environment, as this
compound is released from other organisms. In addition, we have
demonstrated that R. sphaeroides is
able to use 3-hydroxypropionate as a sole carbon source (Schneider,
2011). If 3-hydroxypropionate were exclusively oxidized to
acetyl-CoA and CO2, a functional ethylmalonyl-CoA pathway for
acetyl-CoA assimilation would be essential to convert acetyl-CoA for anaplerosis (see above).
However, inactivation of the gene (ccr) for crotonyl-CoA carboxylase, a neccessary enzyme of the ethylmalonyl-CoA pathway, did not affect growth during 3-hydroxypropionate-dependent growth. Instead, acuI,
encoding acrylyl-CoA reductase (Asao and Alber, 2013), was
shown to be essential for growth with 3-hydroxypropionate and for the
reductive conversion of 3-hydroxypropionate to propionyl-CoA in cell
extracts. Upon production of propionyl-CoA, the cells would be
able to generate succinyl-CoA from which all of the cell's precursor
metabolites can be manufactured (See Figure 4), obviating the need for the ethylmalonyl-CoA pathway.
Results regarding R. sphaeroides growth on 3-hydroxypropionate has prompted us to further investigate the functional role of acrylyl-CoA reductase (AcuI). After working toward characterizing AcuI (Asao and Alber, 2013), it became remarkably curious that the presence of ccr in trans was able to partially restore growth on 3-hydroxypropionate in an acuI null mutant. Additionally, biochemical characterization of AcuI has demonstrated that it forms a new class of acrylyl-CoA reductases (Asao and Alber, 2013), and its amino acid sequence shows similarity to Ccr. In an effort to satisfy our curiousity, we are developing an understanding of the functional characteristics that distinguish AcuI (enoyl-CoA reductase) from Ccr (enoyl-CoA carboxylase/reductase).
Andersen G, et al. 2008. A second pathway to degrade pyrimidine nucleic acid precursors in eukaryotes. J. Mol. Biol. 380: 656–666.
Ansede JH, Pellechia PJ, Yoch DC. 1999. Metabolism of acrylate to β-hydroxypropionate and its role in dimethylsulfoniopropiontae lyase induction by a salt marsh sediment bacterium, Alcaligenes faecalis M3A. Appl. Environ. Microbiol. 65: 5075–5081.
Ansede JH, Pellechia PJ, Yoch DC. 2001. Nuclear magnetic resonance analysis of [1-13C]dimethylsulfoniopropionate (DMSP) and [1-13C]acrylate metabolism by a DMSP lyase-producing marine isolate of the α-subclass of Proteobacteria. Appl. Environ. Microbiol. 67: 3134–3139.
Asao, M. and B. E. Alber. 2013 Acrylyl-CoA reductase, and ezyme in volved in the assimilation of 3-hydroxypropionate by Rhodobacter sphaeroides. J. Bact. In Press.
Berg IA, Kockelkorn D, Buckel W, Fuchs G. 2007. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318: 1782–1786.
Berg IA, et al. 2010. Autotrophic carbon fixation in archaea. Nat. Rev. Microbiol. 8: 447–460.
Callely AG, Lloyd D. 1964. The metabolism of propionate in the colourless alga, Prototheca zopfii. Biochem. J. 92: 338–345
Erb, T. J. et al. (2007) Synthesis of C5-dicarboxylic acids from C2-units involving crotonyl-CoA carboxylase/reductase: The ethymalonyl-CoA pathway. Proc. Nat. Acad. Sci.: 25: 10631 – 10636.
Giovanelli J, Stumpf PK. 1958. Fat metabolism in higher plants. X. Modified β oxidation of propionate by peanut mitochondria. J. Biol. Chem. 231: 411–426.
González JM, Kiene RP, Moran MA. 1999. Transformation of sulfur compounds by an abundant lineage of marine bacteria in the α-subclass of the class Proteobacteria. Appl. Environ. Microbiol. 65: 3810–3819.
Hanarnkar PP, Nelson JH, Heisler CR, Blomquist GJ. 1985. Metabolism of propionate to acetate in the cockroach Periplaneta americana. Arch. Biochem. Biophys. 236: 526–534.
Herter S, et al. 2001. Autotrophic CO2 fixation by Chloroflexus aurantiacus: study of glyoxylate formation and assimilation via the 3-hydroxypropionate cycle. J. Bacteriol. 183: 4305–4316
Kornberg, H. L. and Krebs, H. A. (1957) Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle. Nature: 179: 988 – 991.
Kornberg, H. L. and Lascelles. (1960) The Formation of Isocitratase by the Athiorhodaceae. J. gen. Microbiol: 23: 511-217.
Lloyd D, Evans DA, Venables SE. 1968. Propionate assimilation in the flagellate Polytomella caeca. An inducible mitochondrial enzyme system. Biochem. J. 109: 897–907.
Loh KD, et al. 2006. A previously undescribed pathway for pyrimidine catabolism. Proc. Natl. Acad. Sci. U. S. A. 103: 5114–5119.
Lucas KA, Filley JR, Erb JM, Graybill ER, Hawes JW. 2007. Peroxisomal metabolism of propionic acid and isobutyric acid in plants. J. Biol. Chem. 282: 24980–24989.
Luo LH, et al. 2011. Identification and characterization of the propanediol utilization protein PduP of Lactobacillus reuteri for 3-hydroxypropionic production from glycerol. Appl. Microbiol. Biotechnol. 89: 697–703.
Osterman A. 2006. A hidden pathway exposed. Proc. Natl. Acad. Sci. U. S. A. 103: 5637–5638
Qatibi AI, Bennisse R, Jana M, Garcia J-L. 1998. Anaerobic degradation of glycerol by Desulfovibrio fructosovorans and D. carbinolicus and evidence for glycerol-dependent utilization of 1,2-propanediol. Curr. Microbiol. 36: 283–290.
Rendina G, Coon MJ. 1957. Enzymatic hydrolysis of coenzyme A thiol esters of β-hydroxypropionic and β-hydroxyisobutyric acids. J. Biol. Chem. 225: 523–534.
Schneider, K., M. Asao, M. S. Carter, B. E. Alber. 2012 Rhodobacter sphaeroides uses a reductive route via propionyl coenzyme A to assimilate 3-hydroxypropionate. J. Bact. 194(2): 225.
Strauss G, Fuchs G. 1993. Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle. Eur. J. Biochem. 215: 633–643.
Talarico TL, Casas IA, Cung TC, Dobrogosz WJ. 1988. Production and isolation of reuterin, a growth inhibitor produced by Lactobacillus reuteri. Antimicrob. Agents Chemother. 32: 1854–1858.
Teufel R, Kung JW, Kockelkorn D, Alber BE, Fuchs G. 2009. 3-Hydroxypropionyl-coenzyme A dehydratase and acryloyl-coenzyme A reductase, enzymes of the autotrophic 3-hydroxypropionate/4-hydroxybutyrate cycle in the Sulfolobales. J. Bacteriol. 191: 4572–4581.
Todd JD, et al. 2007. Structural and regulatory genes required to make the gas dimethyl sulfide in bacteria. Science 315: 666–669.
Todd JD, et al. 2010. Molecular dissection of acrylate bacterial catabolism—unexpected links with dimethylsulfoniopropionate catabolism and dimethyl sulfide production. Environ. Microbiol. 12: 327–343.
Wagner C, Stadtman ER. 1962. Bacterial fermentation of dimethyl-β-propiothetin. Arch. Biochem. Biophys. 98: 331–336.
Growth on substrates that are exclusively metabolized via acetyl-CoA requires one of many specialized pathways for anaplerosis.
In
order to understand why anaplerotic reactions are required, one must
examine
the relationship between acetyl-CoA and the tricarboxylic acid (TCA)
cycle (Figure 1). Figure 1 illustrates that acetyl-CoA is the
sole source of carbon input for the TCA cycle while intermediates of
the cycle are necessarily removed for biosynthetic purposes.
In the absence of an ancillary pathway for anaplerosis,
acetyl-CoA, a two-carbon molecule, enters the citric acid cycle
as the sole input of carbon. The cycle outputs two carbons as CO2
while intermediates of the cycle are simultaneously withdrawn as
biosynthetic presursors. Therefore, intermediates required for
maintaining the cycle are removed from without any net carbon
input, collapsing the cycle. Ultimately, the cell needs a method
to
synthesize additional intermediates from acetyl-CoA to continue
production of the biosynthetic precursors within the cycle.The necessity for anaplerotic reactions was immediately apparent to Kornberg and Krebbs in 1957 when th
ey
identified the enzyme activities for isocitrate lyase and malate
synthase in
Pseudomonas
and Escherichia coli
(Kornberg and Krebbs, 1957). Together, these enzymes
provided a method for incorporating an additional acetyl-CoA unit while
bypassing
the CO2 releasing steps of the TCA cycle. They
had seemingly solved the question of acetyl-CoA assimilation.
With the newly
understood "glyoxylate bypass," as they termed the pathway, an organism
could assimilate carbon during growth solely on substrates that are
metabolized via acetyl-CoA. However, they shortly realized a
new complexity. Some organisms, including R. sphaeroides, do
not exhibit isocitrate lyase activity in cell extract but were fully
capable of
assimilating acetyl-CoA (Kornberg and Lacelles, 1960). For
nearly half a century, a hole loomed in the knowledge of
central carbon metabolism. No researcher was able to identify the
pathway that made it possible for these isocitrate lyase-negative
organisms to assimilate acetyl-CoA until 2007. In 2007, a
unique series of enzymatic activites was discovered in R. sphaeroides
cell
extract that
would make it possible for the organism to assimilate acetyl-CoA (Erb
et al., 2007). This series of reactions has come to be known
as the ethylmalonyl-CoA pathway (Figure 3).
At its essence, the ethylmalonyl-CoA pathway incorporates three
acetyl-CoA
units and two CO2
equivalents and produces
succinyl-CoA
and malate, two citric acid cycle
intermediates. The first step is the condensation of two
acetyl-CoA molecules to
form
acetoactyl-CoA. After a series of hydrations, carboxylations,
and carbon rearrangments, the pathway passes through its
namesake, ethylmalonyl-CoA, which is converted to beta-methylmalonyl-CoA.
The latter is then cleaved to form two branches in the pathway.
One branch leads
to malate by incorporation of a third acetyl-CoA unit while the other
branch proceeds through a set of reactions known from
propionate metabolism to form succinyl-CoA. All of this is
done without any loss of carbon to
carbon dioxide. In fact there are actually two carbon
dioxide units
that are co-assimilated. With the enzymatic reaction series elucidated, our next task is to study the regulation of the ethylmalonyl-CoA pathway. We are currently seeking to identify the level of regulation that is primarily used by R. sphaeroides to control the expression of this pathway and which genetic elements and gene products might be involved in regulating the pathway.
The C3 compound 3-hydroxypropionate (CH2OH-CH2-COO−) is increasingly being recognized as an important intermediate or end product of carbon metabolism in a variety of organisms. So far, there are at least five known metabolic processes involving 3-hydroxypropionate.
1. Propionyl-CoA metabolism in plants:
Propionyl-CoA
is derived from the breakdown of chlorophyll, odd-chain fatty acids, or
amino acids like isoleucine and is oxidized to 3-hydroxypropionate
which is then further oxidized to acetyl-CoA (Giovanelli and Stumpf,
1958;
Lucas et al., 2007; Rendina and Coon, 1957). Some animals and algae may
also metabolize propionate via a similar route (Callely and Lloyd 1964;
Hanarnkar et al., 1985; Lloyd et al., 1968).
2. Some autotrophic CO2 fixation pathways:
In
bacteria and archaea, the reductive conversion of acetyl-CoA and CO2 to
propionyl-CoA via 3-hydroxypropionate is part of two CO2 fixation
pathways; however, different enzymes are used in either pathway to
catalyze the common steps in the conversion of acetyl-CoA and CO2 to
propionyl-CoA (Berg et al., 2007; Berg et al., 2002; Herter et al.
2001; Talarico et al., 1988). For example, the reductive conversion of
3-hydroxypropionate to propionyl-CoA is catalyzed by a fusion protein,
named propionyl-CoA synthase, in Chloroflexus aurantiacus (3-hydroxypropionate bi-cycle), whereas Metallosphaera sedula
(hydroxypropionate/4-hydroxybutyrate cycle) requires three separate
enzymes to catalyze the same reaction sequence (Alber and Fuchs, 2002;
Teufel et al., 2009).
3. Dimethylsulfonopropionate (DMSP) metabolism.
3-Hydroxypropionate
is an intermediate in the metabolism of the secondary metabolite
DMSP by microorganisms (Ansede et al., 1999; Todd et al., 2007). DMSP
is synthesized by marine algae and some land plants, and there are
currently three different mechanisms known for the initial step of DMSP
degradation: demethylation to methylmercaptopropionate (Talarico et
al., 1988, Teufel et al., 2009), cleavage by a DMSP lyase into
dimethylsulfide and acrylate (Ansede et al., 1999; Hanarnkar et al.,
1985; Wagner and Stadtman, 1962), and the cleavage of DMSP into
3-hydroxypropionate and dimethylsulfide by an unusual CoA-transferase
(Todd et al., 2007; Todd et al., 2010). Acrylate or
3-hydroxypropionate generated from the cleavage of DMSP may be further
metabolized to acetyl-CoA and CO2; in the case of acrylate, this
proceeds via 3-hydroxypropionate (Ansede, 1999; Ansede 2001; Todd et
al., 2007). However, some bacteria use DMSP solely as a sulfur source
and may therefore release 3-hydroxypropionate or acrylate as an end
product (Gonzalez et al., 1999).
4. Uracil degradation:
3-Hydroxypropionate has been identified as the end product of two different pathways for uracil degradation in bacteria like Escherichia coli as well as in the yeast Saccharomyces kluyveri (Andersen et al, 2008; Loh et al., 2006, Osterman, 2006).
5. Anaerobic metabolism of glycerol:
There
have been reports of 3-hydroxypropionate formation by the fermentation
of glycerol by lactic acid bacteria (Luo et al. 2011; Talarico et al.,
1988) and the anaerobic oxidation of glycerol by a sulfate-reducing
bacterium (Qatibi et al., 1998).
Considering its abundance, 3-hydroxypropionate is likely to play an important role in the overall carbon cycle as an end product or intermediate in the carbon metabolism of a variety of compounds. However, its use as a carbon source in bacteria is poorly understood.
Figure 4
Rhodobacter sphaeroides
likely encounters 3-hydroxypropionate in its environment, as this
compound is released from other organisms. In addition, we have
demonstrated that R. sphaeroides is
able to use 3-hydroxypropionate as a sole carbon source (Schneider,
2011). If 3-hydroxypropionate were exclusively oxidized to
acetyl-CoA and CO2, a functional ethylmalonyl-CoA pathway for
acetyl-CoA assimilation would be essential to convert acetyl-CoA for anaplerosis (see above).
However, inactivation of the gene (ccr) for crotonyl-CoA carboxylase, a neccessary enzyme of the ethylmalonyl-CoA pathway, did not affect growth during 3-hydroxypropionate-dependent growth. Instead, acuI,
encoding acrylyl-CoA reductase (Asao and Alber, 2013), was
shown to be essential for growth with 3-hydroxypropionate and for the
reductive conversion of 3-hydroxypropionate to propionyl-CoA in cell
extracts. Upon production of propionyl-CoA, the cells would be
able to generate succinyl-CoA from which all of the cell's precursor
metabolites can be manufactured (See Figure 4), obviating the need for the ethylmalonyl-CoA pathway.Results regarding R. sphaeroides growth on 3-hydroxypropionate has prompted us to further investigate the functional role of acrylyl-CoA reductase (AcuI). After working toward characterizing AcuI (Asao and Alber, 2013), it became remarkably curious that the presence of ccr in trans was able to partially restore growth on 3-hydroxypropionate in an acuI null mutant. Additionally, biochemical characterization of AcuI has demonstrated that it forms a new class of acrylyl-CoA reductases (Asao and Alber, 2013), and its amino acid sequence shows similarity to Ccr. In an effort to satisfy our curiousity, we are developing an understanding of the functional characteristics that distinguish AcuI (enoyl-CoA reductase) from Ccr (enoyl-CoA carboxylase/reductase).

Andersen G, et al. 2008. A second pathway to degrade pyrimidine nucleic acid precursors in eukaryotes. J. Mol. Biol. 380: 656–666.
Ansede JH, Pellechia PJ, Yoch DC. 1999. Metabolism of acrylate to β-hydroxypropionate and its role in dimethylsulfoniopropiontae lyase induction by a salt marsh sediment bacterium, Alcaligenes faecalis M3A. Appl. Environ. Microbiol. 65: 5075–5081.
Ansede JH, Pellechia PJ, Yoch DC. 2001. Nuclear magnetic resonance analysis of [1-13C]dimethylsulfoniopropionate (DMSP) and [1-13C]acrylate metabolism by a DMSP lyase-producing marine isolate of the α-subclass of Proteobacteria. Appl. Environ. Microbiol. 67: 3134–3139.
Asao, M. and B. E. Alber. 2013 Acrylyl-CoA reductase, and ezyme in volved in the assimilation of 3-hydroxypropionate by Rhodobacter sphaeroides. J. Bact. In Press.
Berg IA, Kockelkorn D, Buckel W, Fuchs G. 2007. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318: 1782–1786.
Berg IA, et al. 2010. Autotrophic carbon fixation in archaea. Nat. Rev. Microbiol. 8: 447–460.
Callely AG, Lloyd D. 1964. The metabolism of propionate in the colourless alga, Prototheca zopfii. Biochem. J. 92: 338–345
Erb, T. J. et al. (2007) Synthesis of C5-dicarboxylic acids from C2-units involving crotonyl-CoA carboxylase/reductase: The ethymalonyl-CoA pathway. Proc. Nat. Acad. Sci.: 25: 10631 – 10636.
Giovanelli J, Stumpf PK. 1958. Fat metabolism in higher plants. X. Modified β oxidation of propionate by peanut mitochondria. J. Biol. Chem. 231: 411–426.
González JM, Kiene RP, Moran MA. 1999. Transformation of sulfur compounds by an abundant lineage of marine bacteria in the α-subclass of the class Proteobacteria. Appl. Environ. Microbiol. 65: 3810–3819.
Hanarnkar PP, Nelson JH, Heisler CR, Blomquist GJ. 1985. Metabolism of propionate to acetate in the cockroach Periplaneta americana. Arch. Biochem. Biophys. 236: 526–534.
Herter S, et al. 2001. Autotrophic CO2 fixation by Chloroflexus aurantiacus: study of glyoxylate formation and assimilation via the 3-hydroxypropionate cycle. J. Bacteriol. 183: 4305–4316
Kornberg, H. L. and Krebs, H. A. (1957) Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle. Nature: 179: 988 – 991.
Kornberg, H. L. and Lascelles. (1960) The Formation of Isocitratase by the Athiorhodaceae. J. gen. Microbiol: 23: 511-217.
Lloyd D, Evans DA, Venables SE. 1968. Propionate assimilation in the flagellate Polytomella caeca. An inducible mitochondrial enzyme system. Biochem. J. 109: 897–907.
Loh KD, et al. 2006. A previously undescribed pathway for pyrimidine catabolism. Proc. Natl. Acad. Sci. U. S. A. 103: 5114–5119.
Lucas KA, Filley JR, Erb JM, Graybill ER, Hawes JW. 2007. Peroxisomal metabolism of propionic acid and isobutyric acid in plants. J. Biol. Chem. 282: 24980–24989.
Luo LH, et al. 2011. Identification and characterization of the propanediol utilization protein PduP of Lactobacillus reuteri for 3-hydroxypropionic production from glycerol. Appl. Microbiol. Biotechnol. 89: 697–703.
Osterman A. 2006. A hidden pathway exposed. Proc. Natl. Acad. Sci. U. S. A. 103: 5637–5638
Qatibi AI, Bennisse R, Jana M, Garcia J-L. 1998. Anaerobic degradation of glycerol by Desulfovibrio fructosovorans and D. carbinolicus and evidence for glycerol-dependent utilization of 1,2-propanediol. Curr. Microbiol. 36: 283–290.
Rendina G, Coon MJ. 1957. Enzymatic hydrolysis of coenzyme A thiol esters of β-hydroxypropionic and β-hydroxyisobutyric acids. J. Biol. Chem. 225: 523–534.
Schneider, K., M. Asao, M. S. Carter, B. E. Alber. 2012 Rhodobacter sphaeroides uses a reductive route via propionyl coenzyme A to assimilate 3-hydroxypropionate. J. Bact. 194(2): 225.
Strauss G, Fuchs G. 1993. Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle. Eur. J. Biochem. 215: 633–643.
Talarico TL, Casas IA, Cung TC, Dobrogosz WJ. 1988. Production and isolation of reuterin, a growth inhibitor produced by Lactobacillus reuteri. Antimicrob. Agents Chemother. 32: 1854–1858.
Teufel R, Kung JW, Kockelkorn D, Alber BE, Fuchs G. 2009. 3-Hydroxypropionyl-coenzyme A dehydratase and acryloyl-coenzyme A reductase, enzymes of the autotrophic 3-hydroxypropionate/4-hydroxybutyrate cycle in the Sulfolobales. J. Bacteriol. 191: 4572–4581.
Todd JD, et al. 2007. Structural and regulatory genes required to make the gas dimethyl sulfide in bacteria. Science 315: 666–669.
Todd JD, et al. 2010. Molecular dissection of acrylate bacterial catabolism—unexpected links with dimethylsulfoniopropionate catabolism and dimethyl sulfide production. Environ. Microbiol. 12: 327–343.
Wagner C, Stadtman ER. 1962. Bacterial fermentation of dimethyl-β-propiothetin. Arch. Biochem. Biophys. 98: 331–336.